A while ago I wrote a post asking whether everyone’s favourite measure of biodiversity, species richness, was useful. In it, I concluded that it is probably one of the bluntest, least informative measures of ecological communities we have and that we should try to use alternative metrics when possible. Recently, I started wondering about what other measures of biodiversity might be informative, and what they can be used for. And then a neat review of beta-diversity by James Jacob Socolar ( correction courtesy of James Gilroy on Twitter – thanks James!) and colleagues came out in Trends in Ecology and Evolution, so today I’ll focus on that, borrowing from some of their thoughts and hopefully adding some of my own along the way. In the future, at some point, I’ll write something about temporal changes in ecological communities at individual sites.
So, firstly what do I mean by beta-diversity? Beta-diversity broadly reflects the differences in community composition between sites. Gamma diversity (regional diversity) is a product of both beta- and alpha-diversity (diversity at a single site). And there are lots* of different ways of measuring beta-diversity. The simplest metric for beta-diversity is termed ‘true beta-diversity’ and was defined by Whittaker in 1960 as:

This metric is perhaps the easiest to interpret, but it also needs a reliable estimate of gamma diversity, so may be difficult to use in practice. Using this method allows the relationship between alpha and gamma diversity to be investigated. Other measures can be based on dissimilarity matrices, identifying pairwise differences between sites. These metrics can then be used to look at drivers of these differences, such as the geographic distance between individual sites and environmental differences. However, dissimilarity matrix methods don’t allow the relationship between alpha and gamma diversity to be investigated. The above explanation probably explains the ubiquity of species richness as a metric in ecology – we can all (more-or-less) agree on what it means.
Changes in beta-diversity when humans alter natural landscapes can be unpredictable. When human disturbances are patchy, such as in the case of selective logging, beta diversity has been shown to be stable or increase due to an influx of generalist species in forest gaps.
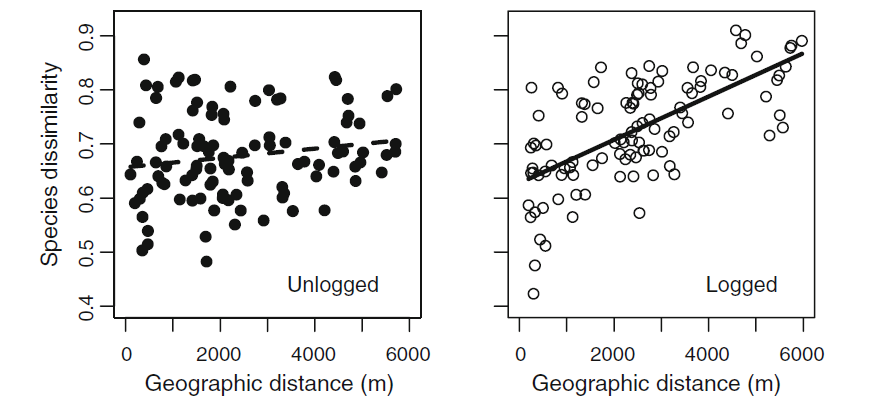
In contrast, when human land-use change results in the conversion of natural ecosystems to a relatively homogeneous system in which only a small subset of species can survive, beta-diversity tends to decrease. Examples of such drivers include agricultural conversion and urbanisation. However, even high intensity farming can result in an increase in beta-diversity particularly if species populations decrease leading to greater dissimilarity purely as a result of random processes. In summary, the response of beta-diversity to anthropogenic change appears to be relatively idiosyncratic.
All of this is well and good, but what use is beta-diversity to practical conservation? At first inspection, this is not clear. The general perception of species richness is that more species = better**. Does higher beta-diversity = better? Well, no, not necessarily. Given that the aims of conservation vary from place to place, it is not surprising that how beta-diversity can be used also varies.
The most obvious use of beta-diversity is in spatial planning of protected areas. In landscapes which show a high spatial turnover of species, managers might favour the use numerous distinct reserves to capture this variation. However, in a landscape in which beta-diversity results from differents in species richness a single protected area might be favoured. Also, if a natural ecosystem is particularly distinct from other candidate sites it may be considered a priority for protection.
High beta-diversity can also result from dispersal limitation in a landscape. For example, secondary forests in fragmented landscapes plants with seeds dispersed by wind may colonise sites more readily than those dispersed by animals that may not cross non-forest areas. So in cases where beta-diversity amongst patches of a similar habitat in a fragmented landscape is high, this may point to the need for restoration to increase connectivity. Successful restoration may result in a decrease of beta-diversity as dispersal between patches increases. For example, Renata Pardini’s work has shown that the small mammal communities of more highly connected fragments of Atlantic forest are more similar to other patches than unconnected fragments. However, as far as I know, there is relatively little evidence empirical that restoration has similar effects.
In the paper I mentioned earlier, Jacob Socolar and colleagues suggest that beta-diversity may also be useful in informing the land-sharing vs land-sparing debate (which i have previously written about here, here an here). They argue that the use of beta-diversity as part of this debate may show that heterogenous landscapes that include agri-environment schemes, management of natural systems and high intensity agriculture are better at maintaining alpha- beta and gamma-diversity. Thus, the incorporation of metrics other than population sizes of species, the classic approach for such comparisons, may produce different conclusions to current studies, which largely suggest land-sparing as a favoured approach. As always with conservation, this depends on what you think we should try to protect. Should we focus on particular species? Or should we look attempt to conserve the processes that maintain coarse-scale diversity?
For me, the key point that the paper makes is that even though two recent high-profie studies recently suggested local-scale alpha-diversity is relatively constant***, global scale gamma-diversity is declining. This suggests that rare species are getting rarer and common species are increasing in abundance. If we can work out how and why beta-diversity responds to land-use changes we can better understand how to conserve gamma-diversity. However, before we do that we need to develop methods to upscale from alpha to gamma diversity and determine how different disturbances alter beta-diversity. Novel approaches offer the potential to solve this problem, but substantial testing is needed to determine how useful they are.
*Patricia Koleff identified 24 metrics for use with presence-absence data and my old CEH office mate Louise Barwell tested 29 different beta-diversity metrics that incorporated abundance data. Give both of these papers a read, they’re well worth your time.
**I don’t agree with this perception, I’m just extrapolating based on things I have heard from a few people. Deeply unscientific, I know.
***I saw Andrew Gonzalez present some work on the problems of these two studies at the 2015 British Ecological Society annual meeting and hope to post something when the paper comes out. I can’t say much, but it was fascinating stuff.


